Neurons

































Introduction
Neurons are the building block cells of our central nervous system, especially in our brain. Although there are many different types of neurons, as illustrated in the drawing at left by Santiago Ramón y Cajal, almost all of them share some basic structural elements and functionality. We’ll explore these commonalities in this activity.
Instructions
In all sections except the one on Neuron Structure, simply click on the images in the left portion of the screen to toggle between them (image numbers referred to in the explanations will appear in the upper right of the yellow area of the screen).
The first time you do this activity, you should probably go through the parts in order from top to bottom. In the Neuron Structure section, click on each part of the neuron to see its name and read what it does.
Neuron Structure
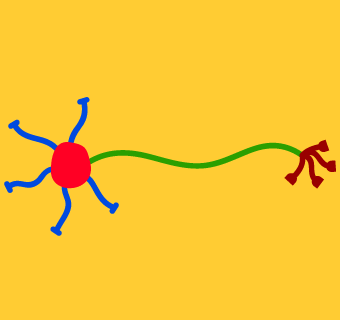
The image at left shows a cartoon of a simplified neuron. Click on each different-colored part of the neuron to show its name and read about its function.
In this portion of the activity we will simplify things by talking about how information flows through the various parts of a neuron. In subsequent sections, we’ll get into how this information is actually communicated.
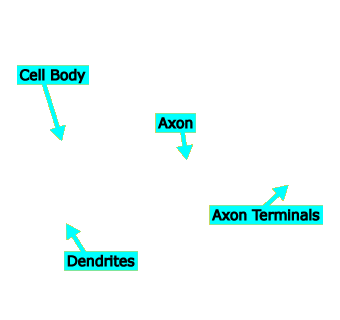
Cell Body
Like all cells, every neuron has a cell body, sometimes called the soma. The cell body contains the nucleus, which holds the cell’s chromosomes, and is responsible for performing the basic metabolic activities that help keep the cell alive.
In most neurons, the cell body also serves as a kind of collection center for information coming in through the dendrites from upstream neurons. If the cell body is excited enough about the information it receives from the dendrites, it will pass a signal down the axon, through the axon terminals, and across synapses to downstream neurons.
Axon
Almost all neurons have a single axon, which transmits neural information from the cell body, through the axon terminals, and finally across synapses to other neurons. Axons can be very long—up to several meters for neurons in giraffes’ necks.
Axon Terminals
Although almost all neurons have only a single axon, most axons branch out at their ends into a number of axon terminals. Each axon terminal synapses with one or more downstream neurons. Thus, every neuron can “talk to” a large number of subsequent neurons.
Dendrites
Dendrites receive information from upstream neurons and transmit it to the cell body. A neuron may have tens, hundreds, or even thousands of dendrites, so each neuron “listens to” a large number of preceding neurons.
Information Flow Through a Neuron: The Action Potential
In the previous part of the activity we saw how information flows into a neuron through its dendrites, then passes through the cell body, down the axon, and out to other neurons via axon terminals. We also saw that most of this journey actually takes place in the axon, which is typically much longer than the dendrites or cell body.
Here, we examine more closely how information passes through the axon. Much of what we know about this process was learned through the sacrifice of squids (Image 1), which possess neurons with particularly thick—and therefore relatively easy to study—axons.
In essence, an axon is a long tube formed by the neuron’s cell membrane, with cytoplasm (the jelly-like material that fills cells) on the inside and interstitial fluid (the water-like liquid in which cells are bathed) on the outside (Image 2). Because there are normally more negatively charged particles (relative to positively charged particles) in the cytoplasm than in the interstitial fluid, the inside of the axon normally has a small, negative electrical potential.
Suppose we extract a giant squid neuron (that’s a giant neuron from a squid, not a neuron from a giant squid), keep it alive in a Petri dish, and insert a small electrode and an electrical potential recorder at the points indicated in Image 3. As shown in the graph in the upper right corner of the image, we would at first measure a negative electrical potential, for the reasons noted above.
Now suppose we apply a small electrical charge through the electrode (Image 4). As shown in the graph, this will lead to a small bump in electrical activity for a brief moment, but nothing any more spectacular than this will happen. Applying a slightly larger charge (Image 5) produces a slightly bigger bump.
If we increase the charge a bit more, though, something interesting happens (Image 6): we record a sudden and brief spike in electrical activity, greater than the initial charge we applied. This spike is called an action potential. As shown in Image 7, applying an even larger initial charge does not lead to a larger action potential. Thus, we see that the action potential is an “all-or-nothing” phenomenon. It is either generated or not, depending on whether the initial charge is big enough, and the size of the action potential is fixed.
Two other interesting features of axon potentials are illustrated in Image 8. First, action potentials propagate down the length of axons. That is, once an action potential is initiated, the spike of electrical potential travels in a wave down the length of the axon. Second, this propagation process only goes in one direction: the action potential is always generated at the cell body and always propagates down the length of the axon to the axon terminal.
Once the action potential reaches the axon terminals, neural information is communicated to other neurons across synapses, as explained in the next part of the activity.
Information Flow Between Neurons: Synapses and Neurotransmitters
Although neurons were once thought to touch each other and thereby pass electrical signals directly, we now know that a small gap, called a synapse, separates the axon terminal of one neuron from the dendrite of another. (The debt for our knowledge of the workings of synapses is owed largely to the lowly Aplysia, better known as the sea slug; Image 1.)
Image 2 shows our cartoon neuron from part 1 of the activity, with its dendrites receiving information from axon terminals of upstream neurons across five synapses, and its axon terminals sending information to dendrites of downstream neurons across four more synapses.
Image 3 shows a close-up of one of these synapses. The presynaptic (upstream) neuron’s axon terminal is shown in maroon, and the postsynaptic (downstream) neuron’s dendrite in blue.
When an action potential reaches the axon terminal, small “bubbles” called synaptic vesicles start moving through the axon terminal’s cytoplasm toward the cell membrane (Image 4). When they reach the membrane, the synaptic vesicles release their contents—chemicals called neurotransmitters—into the synaptic cleft between the neurons. The neurotransmitters then diffuse across the synaptic cleft and are taken up by receptors embedded in the postsynaptic dendrite’s cell membrane (Image 5).
What happens next depends on a number of factors. Some neurotransmitters are excitatory, meaning that when they enter the postsynaptic neuron’s dendrite, they make that neuron more likely to fire an action potential. However, other neurotransmitters are inhibitory, meaning that they try to prevent the postsynaptic neuron from firing an action potential. The interplay between excitatory and inhibitory synaptic connections is explored in the next part of this activity.
Excitation and Inhibition
Image 1 shows our cartoon neuron, labeled here as “Neuron 0,” receiving information from five different presynaptic axon terminals (neurons 1–5). Let us suppose that three of the presynaptic axon terminals release excitatory neurotransmitters (as indicated by the green circles), whereas the other two release inhibitory neurotransmitters (red circles).
If an action potential fires down the axon of neuron 1 (Image 2), the subsequent excitatory neurotransmitter release causes neuron 0 to fire an action potential.
However, suppose now that neurons 1 and 3 both fire at the same time (Image 3). The excitation generated by neuron 1’s neurotransmitters is cancelled out by the inhibition generated by neuron 3’s neurotransmitters. The net result is that neuron 0 does not fire.
If neurons 1, 3, and 4 all fire at once (Image 4), we would have two sources of excitation and only one source of inhibition. Therefore, neuron 0 would once again fire since the excitatory input would outweigh the inhibitory input.
This is an oversimplification of the complicated workings of neurons and neurotransmitters, but the general point is still valid. Whether or not neuron 0 fires depends on the sum of the inhibitory and excitatory inputs from neurons 1–5. The summation of inputs occurs in the neuron’s cell body.
One question you might be asking yourself at this point is what happens if neurons 1, 3, and 4 aren’t firing at exactly the same time. The answer to this question is that it doesn’t really matter, because what matters with neurons is not really single action potentials, but action potential rates. We’ll explore this concept in the next part of the activity.
Neural Firing Rates
In the earlier section on action potentials we showed what happens if successively larger electrical shocks are applied to a neuron. We saw that if the shock is powerful enough, an action potential is generated (Image 1).
Other experiments have shown that neurons actually fire action potentials spontaneously. In other words, if we measure the electrical potential of a neuron without applying any external electrical stimulation, the neuron will still fire a signal down its axon on its own every now and then. The frequency with which a neuron fires such spontaneous action potentials is called its resting rate.
We can represent neural firing rates with graphs like the one shown in Image 2. Here, each vertical line or “spike” on the graph indicates a point in time when the neuron fired an action potential.
There are a few special types of neurons that have no dendrites. These are called sensory receptors, and their job is to detect various types of energy in the environment. In other words, receptors are responsible for starting the whole process of sensation and perception. There are several types of receptors lodged in your skin, whose job it is to detect mechanical pressure, temperature changes, tissue damage, or other touch sensations.
We’ll explore these particular receptors in detail in Chapter 13. For now, though, let’s consider what would happen if we recorded the action potentials generated by a touch receptor in your finger as you tap it on your desk. As we just noted, even if you weren’t tapping your finger at all, the receptor would fire spontaneously at its resting rate (Image 2).
If you were to tap your finger lightly on the desktop, the receptor’s firing rate would increase slightly (Image 3). Tap harder and harder, and the rate would increase more (Image 4) and more (Image 5).
At some point, however, the receptor’s firing rate would max out. If you mash your finger down as hard as you can, your pain receptors might start firing, but the touch receptor’s firing rate would not increase any more. This limit on neural firing rates is due to the small “dip” in electrical potential that you might have noticed on the right side of our original graph of the action potential (Image 1). During this short recovery period, called the refractory period, the neuron cannot fire again.
In future chapters, you will often read that a receptor or other neuron becomes more active in one situation than in another. For example, we will see that there are two types of visual receptors—rods and cones—and that rods are more active than cones under low light conditions. This does not mean that rods have larger action potentials than cones in the dark. As we saw earlier in this activity, single action potentials are an all-or-nothing phenomenon. They either do or do not occur, and when they occur they always have the same intensity. Instead, greater activation should always be taken to mean a higher rate of firing action potentials.
Neural Networks
Suppose we start with a single neuron that has five dendrites and five axon terminals (Image 1). This neuron receives information from five upstream neurons and passes the information on to five downstream neurons. Now suppose each of this neuron’s dendrites synapses with the axon of another neuron with five dendrites and five axon terminals (Image 2). Then suppose each of the axon terminals of these five neurons synapses with another five-dendrite–five-axon-terminal neuron (Image 3). At this point we’ve created a neural network of 31 cells. Our next layer (Image 4) would add 150 neurons to the network, and the layer after that would add 750 more! Remember that we’re only considering the connections coming in from one of each neuron’s dendrites and that each one is also receiving information from four other neurons.
Now consider that neurons in the human brain might receive information from hundreds of upstream neurons and send information on to hundreds of downstream neurons. Furthermore, real neural networks are massively interconnected. For example, the five neurons in layer 2 of our simplified network might each talk to the others (Image 5). Also, later layers might form feedback loops with earlier layers. And, perhaps most importantly of all, some neural connections are strongly excitatory and others strongly inhibitory, with an infinite number of shades of excitation and inhibition in between.
Put all these factors together and you start to see why the largest neural network of all, the whole human brain (which has something on the order of 100 billion neurons that form 100 trillion synapses), has been called the most complex structure in the known universe.